Spiking Neural Networks (SNNs) are biologically inspired models that mimic the temporal information processing of biological neurons through discrete spikes (Vreeken, 2003). Unlike the first two generations of artificial neural networks, SNNs encode information in the timing of spikes rather than in average firing rates alone (Taherkhani et al., 2019; Ponulak & Kasinski, 2011). In this guide we explore the biological foundations of spiking neural networks, from the biophysics of a single biological neuron to the metastable behaviour of the human brain.
Biological Brains
Recent studies using advanced cell-counting techniques, such as the isotropic fractionator method, estimate that the average adult human brain contains approximately 86 billion neurons (Herculano-Houzel, 2009). In addition to this vast number of neurons, the human brain also contains roughly the same number of non-neuronal cells, such as the glial cells that provide support and protection for neurons (Herculano-Houzel, 2012).
In terms of mass the human brain weighs around 1.5 kg, with 82% of this mass belonging to the outer part of the brain known as the cerebral cortex. Neuron count tells a different story, however: the cerebral cortex holds only around 19% of the brain's neurons (Herculano-Houzel, 2012).
The key to the brain's complexity lies in its interconnectedness, with each of these 86 billion neurons typically connected to more than 10,000 postsynaptic neurons on average (Gerstner & Kistler, 2002). This complexity is far from homogeneous: the brain has specialised regions with different functions and evolutionary histories. The most complex, and relatively unique to developed brains, is the cerebral cortex, where a neuron may make on the order of tens of thousands of synaptic contacts.
Another important region is the cerebellum, responsible for motor control and coordination. The cerebellum contains around 69 billion neurons (more than the cerebral cortex) despite being much smaller in mass (Herculano-Houzel, 2009). It is also unusual in housing Purkinje cells, some of the largest and most elaborately branched neurons in the brain.
A further remarkable property of the brain is its energy profile. The human brain consumes around 20% of the body's total energy budget, an enormous share for a single organ, suggesting that evolution has pushed hard against the trade-off between intelligence and energetic cost. Yet it is remarkably frugal compared with modern computing hardware: at around 20 W, the brain uses far less energy than large language models such as GPT-3, which required over 1,000 MW-hours to train, around 50 million hours of brain-equivalent energy (Zador, 2022).
Biological Neurons
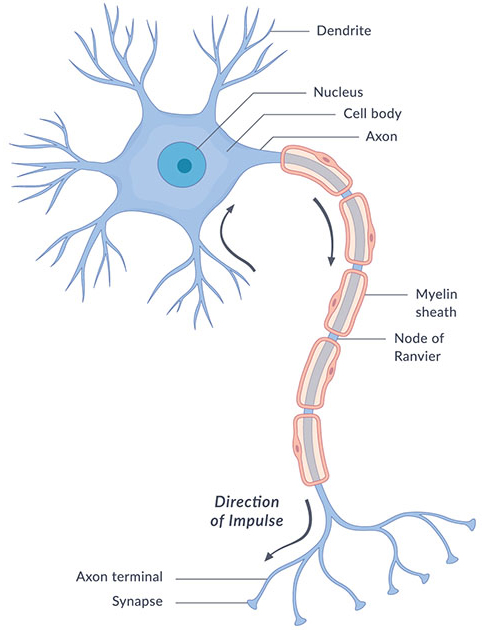
A neuron is organised into four functional regions. The dendrites are branched processes that receive inputs from other cells; the soma, or cell body, houses the nucleus and integrates those inputs; a single axon carries the output away; and the synaptic terminals at the axon's end pass signals on to the next cells. Signal flow is directional: dendrites and soma integrate incoming signals, and if their sum crosses a threshold, a spike is launched down the axon to the terminals (Gerstner et al., 2014).
The neuron maintains a voltage difference across its membrane. At rest the interior sits at roughly −70 mV relative to the outside, a potential set mainly by the membrane's selective permeability to potassium and by the sodium–potassium pump that continuously trades ions across it. Inputs nudge this potential up or down; when it reaches a threshold of about −55 mV, the neuron fires (Betts et al., Anatomy & Physiology). Many axons are wrapped in myelin, a fatty insulating sheath interrupted by gaps called nodes of Ranvier, which lets the signal leap from node to node and dramatically speeds its conduction.
Action Potentials
The spike, or action potential, is an all-or-nothing event: once the membrane crosses threshold, the neuron fires at full amplitude regardless of how much further the input pushed it. The mechanism is a rapid, self-reinforcing dance of voltage-gated ion channels. In depolarisation, sodium channels open and sodium ions rush inward, driving the potential sharply upward toward +30–40 mV. In repolarisation, the sodium channels inactivate while potassium channels open and potassium flows out, pulling the voltage back down. A brief hyperpolarisation overshoots below the resting level before the membrane settles (Betts et al., Anatomy & Physiology).
Immediately after a spike the neuron cannot fire again: during the absolute refractory period the sodium channels are inactivated, and during the following relative refractory period a stronger-than-usual input is needed. This refractoriness caps the maximum firing rate at a few hundred hertz. Conduction speed ranges from around 0.5–2 m/s in unmyelinated axons to over 100 m/s in myelinated ones. The quantitative theory of all this dates to Hodgkin & Huxley (1952), whose conductance-based model of the squid giant axon reproduced the spike's shape and propagation and earned a Nobel Prize, and it remains the most biophysically faithful of the neuron models used in simulation today.
Synaptic Transmission
Neurons communicate across synapses. At the majority (chemical synapses) an arriving spike triggers the release of neurotransmitter molecules that cross a narrow cleft and bind receptors on the receiving cell; a minority are electrical synapses, or gap junctions, that pass current directly for very fast, bidirectional coupling. The effect on the receiving neuron can be excitatory, producing a depolarising post-synaptic potential that pushes it toward threshold, or inhibitory, producing a hyperpolarising one that pulls it away. A neuron's behaviour reflects the running balance of the two.
Because a single input is usually far too small to trigger a spike on its own, neurons perform synaptic integration, combining many inputs by temporal summation (inputs arriving close together in time) and spatial summation (inputs arriving at different points on the dendritic tree). Synapses are also plastic: the strength of a connection changes with use. Donald Hebb's 1949 postulate (that a synapse strengthens when the presynaptic cell reliably helps fire the postsynaptic one, often paraphrased as "cells that fire together wire together") is the conceptual root of learning in these systems. Its precise, biologically observed form is spike-timing-dependent plasticity: Bi & Poo (1998) showed that when a presynaptic spike precedes a postsynaptic one within about 20 ms the synapse is strengthened, whereas the reverse order weakens it, making the relative timing of spikes the learning signal. This rule reappears as a training method for artificial spiking networks under learning approaches.
Neural Coding
How does a train of identical spikes carry information? Under rate coding, what matters is the firing rate: the number of spikes in a window, or the fraction of a population active at an instant. Rate codes are robust to noise but comparatively slow. Under temporal coding, the precise timing carries the message: in time-to-first-spike coding the latency of the earliest spike encodes stimulus strength, while in phase coding the timing relative to an ongoing brain rhythm is what counts. Gerstner et al. (2014) note that pure rate codes can be too slow to explain the speed of biological reaction times, which is one motivation for timing-based codes.
Two further principles matter. In population coding, a stimulus is represented across the joint activity of many neurons with overlapping tuning, giving accuracy and fault tolerance beyond any single cell. In sparse coding, only a small fraction of neurons is active at any moment; Olshausen & Field (1996) famously showed that learning a sparse code for natural images spontaneously produces the localised, oriented, edge-detecting receptive fields seen in primary visual cortex. These codes (precise timing, sparsity, population representation) are exactly what spiking networks are positioned to exploit, and what rate-only artificial networks discard.
Collective Behaviour in the Brain
Intelligence emerges not from single neurons but from their collective dynamics. Populations of neurons fire rhythmically in characteristic frequency bands (delta, theta, alpha, beta and gamma), and Buzsáki & Draguhn (2004) argue that these oscillations form a hierarchical family that coordinates communication across brain regions, binding distributed neurons into transient assemblies and nesting fast rhythms within slower ones.
Strikingly, the resting cortex appears to hover near a critical point. Beggs & Plenz (2003) found that spontaneous activity organises into cascades ("neuronal avalanches") whose sizes follow a power law with an exponent near −3/2, the statistical signature of a system balanced between order and chaos, a regime thought to maximise dynamic range and information transmission. The brain never settles into a fixed state nor fully synchronises; instead it lingers briefly near coordinated configurations and spontaneously switches between them. This metastability, described by Tognoli & Kelso (2014), blends integration and segregation and is thought to be the dynamical regime that makes flexible, ever-shifting cognition possible.
These are the biological phenomena that spiking neural networks aim to borrow from. The rest of this background explains how they are captured in practice: the neuron models that formalise the spike, the network architectures that wire units together, the learning approaches that tune them, the hardware and software that run them, and the applications where they are beginning to pay off.